LA FAUNE
DE
VERTEBRES CORSE
Paul FRANCESCHI
ADECEC-CERVIONI 1994
Avec plus de 8700 km2 la Corse est par sa superficie
la quatrième des grandes îles de la Méditerranée.
Son altitude élevée (plus de 500m en moyenne avec 2710m
à son point culminant) atténue le caractère méditerranéen
du climat et explique en partie la variété des biotopes
rencontrés.
La faune insulaire sera donc, dans son ensemble, le reflet de cette
diversité géographique, tout en se caractérisant
par quelques points particuliers.
Nous nous intéresserons principalement ici aux Vertébrés,
c'est-à-dire aux animaux qu'on a coutume de qualifier de supérieurs.
Cet embranchement animal comprend les Poissons, les Amphibiens (souvent
appelés Batraciens), les Reptiles, les Oiseaux et les Mammifères.
Nous ne parlerons pas des poissons marins qui peuplent les côtes
de la Corse(1), ni des oiseaux migrateurs ou des mammifères volants
(Chauves-souris). Ces groupes possèdent en effet le point commun
de pouvoir se déplacer sur de grandes distances : cette faculté
de déplacement rend difficile la mise en place des mécanismes
d'isolement et donc les phénomènes de spéciation
(cf. note10) : leur intérêt est donc réduit quant
à une contribution originale à la composition faunistique
insulaire.
La faune corse, même si cela semble une évidence, se caractérisera
donc essentiellement par son insularité : barrière ou
filtre (plus ou moins sélectif), protection ou refuge, le fait
géographique est incontournable. Il faut y ajouter, mais dans
une moindre mesure, les contraintes climatiques méditerranéennes
et (en ce qui concerne les mammifères non-volants) l'intervention
humaine, volontaire ou pas.
Il existe plusieurs moyens susceptibles de permettre
la caractérisation des peuplements animaux. L'un des plus simples
consiste à apprécier la richesse de la faune en se référant
au nombre d'espèces présentes dans une aire géographique
donnée.
Si on considère ce premier paramètre la faune de la
Corse en Vertébrés se caractérise, comme toutes
celles des autres îles, par un nombre d'espèces moins
élevé que celui qu'on pourrait trouver dans des régions
continentales comparables (tableau 1).
| Corse | Sardaigne | Midi France | |
| Amphibiens(1) | 7 | 9 | 12 |
| Reptiles | 11 | 15 | 21 |
| Oiseaux | 138(2) | 141(2) | 173(2) |
| Mammifères(3) | 17 | 18 | 45 |
Tableau 1. Comparaison de la richesse en Vertébrés de la Corse et d'autres régions méditerranéennes.
(1)D' après DELAUGERRE et CHEYLAN (1992)
(2)D' après BLONDEL (1986)
(3)D'après NOBLET et al. (1987)
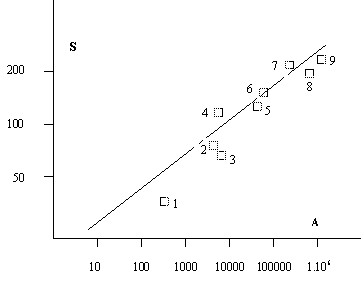
1: Malte; 2: Crête; 3: Chypre; 4: Corse; 5: Sardaigne;
6: Sicile; 7: Balkans; 8: Maghreb; 9: Ibérie + France + Italie
A: superficie (Km2)
S: nombres d'espèces nicheuses
Figure 1: Relation log - log entre nombre d'oiseaux nicheurs et superficie (BLONDEL, 1982)
Le nombre d'espèces est également corrélé avec la superficie de l'aire zoogéographique considérée (figure 1). L'existence de ces deux contraintes (insularité et superficie) permet d'expliquer l'essentiel des variations observées dans la composition des diverses faunes insulaires méditerranéennes (2).
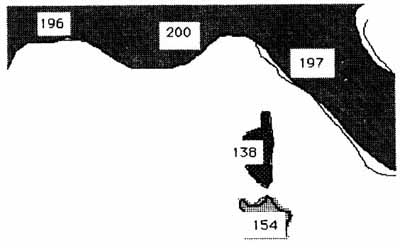
Figure 2 . Nombre d'espèces d'oiseaux nicheurs dans différentes régions du bassin méditerranéen occidental (d'après BLONDEL, 1986)
Cet appauvrissement est général : il touche
aussi bien les oiseaux (figure2) que les Mammifères (17 espèces
contre plus de 40 sur le continent), les Amphibiens et les Reptiles.
Même les poissons d'eau douce sont sujets à ce phénomène
: si l'on exclut les introductions récentes, on compte seulement
4 espèces autochtones dans les cours d'eau de l'île (3).
Les raisons de la réduction du nombre des espèces insulaires
sont multiples et leur importance relative encore discutée. La
plus évidente semble être l'impossibilité, pour
une espèce donnée, de coloniser une aire géographique
à laquelle elle est par trop étrangère, et donc
inadaptée. C'est le cas, en ce qui concerne la Corse, des espèces
arctiques ou boréales inféodées aux forêts
ou aux toundras nordiques.
On peut également envisager, pour un certain nombre d'animaux,
la difficulté voire l'impossibilité à se déplacer
sur de longues distances, comme par exemple franchir des bras de mer
plus ou moins importants. Mais, en fait, il semble bien que dans la
plupart des cas le manque de biotopes favorables et l'importance de
la compétition interspécifique expliquent le mieux l'absence
d'un grand nombre d'espèces dans les îles.
Il faut remarquer à ce sujet que l'appauvrissement de la faune
corse touche inégalement les différents groupes qui la
composent : les insectivores aériens (Hirondelles et Martinets
chez les Oiseaux, Chauves-souris chez les Mammifères) sont ainsi
particulièrement bien représentés, de même
que certains genres de petits passereaux, comme les Fauvettes (4).
La réduction du nombre d'espèces s'accompagne également
de modifications importantes dans les densités de peuplement
(c'est à dire l'occupation de l'espace par les animaux). Mais
si celles-ci sont plus importantes que dans les biotopes continentaux
la taille et le poids des individus sont en général plus
faible.
Les niches écologiques (5) sont le plus souvent modifiées,
en particulier pour ce qui touche au régime et au comportement
alimentaires : on pourrait dire, de manière simplifiée,
que les niches des espèces manquantes sont investies par celles
qui sont là (6). Cette tendance marquée à occuper
l'ensemble de l'espace et des ressources disponibles débouche
évidemment sur un obstacle supplémentaire, d'ordre compétitif,
à l'installation de nouvelles espèces.
Une autre manière de s'intéresser à
la spécificité d'un peuplement animal est de prendre en
compte son degré d'originalité en analysant ses composantes
endémiques.
Les espèces dites endémiques sont caractérisées
par une aire géographique restreinte : nous entendrons ici
la Corse, ou la Corse et la Sardaigne (on parlera d'endémiques
cyrno-sardes) auxquelles on adjoint, pour un certain nombre de cas,
l'archipel toscan et les îles de la côte méditerranéenne
française.
| Nb d'espèces | Endémiques et cyrno-sardes* | Sous espèces | |
| Amphibiens | 7 | 2-1* | 2 (1) |
| Reptiles | 11 | 3 | 1 (2) |
| Oiseaux | 127 | 1-1* | 12 (3) |
| Mammifères | 17 | 1 | 7 (4) |
Tableau 2 . Spécificité de la faune de Vertébrés de la Corse
(1)Les deux sous-espèces concernées (la
rainette et la salamandre) sont parfois considérées comme
des espèces à part entière.
(2)La couleuvre à collier Corse, sans doute identique à
celle de Sardaigne (Natrix natrix cettii) pourrait en fait s'avérer
posséder le statut d'espèce.
(3)Ce nombre de sous-espèces (cyrno-sardes pour la plupart) est
sujet à caution. Pour une revue exhaustive de cette question
voir THIBAULT (1983).
(4)Il s'agit du renard, du chat sauvage, du loir, du lièvre,
du sanglier, du cerf et de la belette.
Si le taux d'endémisme est relativement élevé
chez les Végétaux supérieurs et surtout chez certains
groupes d'Invertébrés (7), il l'est beaucoup moins chez
les vertébrés. Cette différence est plus ou moins
marquée selon les classes considérées (cf. tableau
2). Les Mammifères et les Oiseaux présentent peu d'espèces
endémiques (cf. note 8) par rapport aux Reptiles et surtout aux
Amphibiens (5 espèces sur 7).
Chez les poissons d'eau douce, la sous-espèce corse de truite
(Salmo trutta macrostigma) présente des caractéristiques
génétiques extrêmement originales qui l'éloignent
fortement des populations continentales (9). Il semble clair ici que
l'isolement géographique, ou plus exactement l'importance de
sa durée, a joué un rôle prépondérant
: isolées depuis longtemps de leurs homologues continentales,
certaines populations insulaires peuvent évoluer de façon
divergente, parfois jusqu'à l'acquisition d'un statut de nouvelle
espèce. D'autres, sans doute isolées depuis peu de temps,
se sont moins nettement individualisées et forment aujourd'hui
des sous-espèces ou des espèces vicariantes (10).
La liste des espèces animales propres à la Corse est
réduite à deux Amphibiens (l'euprocte de Corse, Euproctus
montanus, et le discoglosse Corse, Discoglossus montalentii) et à
un Oiseau (la sittelle Corse, Sittella Whitheheadi). On peut y joindre
six espèces cyrno-sardes : un Amphibien (le discoglosse sarde,
Discoglossus sardus), trois Reptiles (le lézard tiliguerta,
Podarcis tiliguerta, le lézard de Bedriaga, Archaeolacerta
bedriagae, et l'algyroïde de Fitzinger, Algyroides fitzingeri),
un Oiseau (le venturon corse, Serinus corsicana) et un Mammifère
(le mouflon de Corse, Ovis ammon musimon).
Si l'on excepte le mouflon on constate qu'il s'agit d'espèces
peu spectaculaires et, pour la plupart, difficiles à observer
: cette liste exclut notamment les grands rapaces encore présents
dans l'île et les espèces les plus courantes d'Oiseaux
et de Mammifères.
En fait, il semble que le Bassin méditerranéen, s'il
a joué un rôle de refuge important lors des glaciations
successives, s'est trouvé moins impliqué dans la genèse
des espèces d'Oiseaux et de Mammifères que dans celle
des Reptiles et des Amphibiens. Ainsi d'après Blondel (1986),
seuls les genres Alectoris (Perdrix), Sitta (Sittelles) et Sylvia
(Fauvettes) semblent renfermer des espèces typiquement méditerranéennes
chez les Oiseaux.
La répartition circum-méditerranéenne des Perdrix
et des Fauvettes est fort instructive. En dehors du nombre relativement
élevé d'espèces présentes chez ces deux
groupes d'oiseaux, il est intéressant de constater que si les
Fauvettes méditerranéennes peuvent cohabiter sans problème,
les Perdrix, elles, s'excluent mutuellement et s'hybrident dans les
régions où elles se rencontrent (11). Cette situation
s'explique certainement par le fait que ces oiseaux, d'ailleurs morphologiquement
très proches, possèdent des niches écologiques
quasi-identiques et ne peuvent subsister ensemble sur un même
territoire : une espèce éliminera toujours l'autre (12).
La présence de zones d'hybridation tendrait à accréditer
la thèse de l'apparition "récente" de ces
espèces et la mise en place incomplète des mécanismes
d'isolement.
Les faunes insulaires, et donc la faune Corse, constituent ainsi de
véritables laboratoires pour tout ce qui touche aux problèmes
évolutifs et aux modèles de spéciation.
L'origine de la plupart des Vertébrés
de Corse est encore discutée. Si l'ancienneté de nombreux
Amphibiens et Reptiles est certaine, celle des Oiseaux et surtout des
Mammifères est plus discutable. En ce qui concerne les Mammifères,
sur lesquels nous nous pencherons plus particulièrement ici,
l'histoire des peuplements insulaires est assez mal connue en ce qui
concerne la plupart des espèces de la faune actuelle.
Un fait semble incontestable : la faune corse pléistocène,
profondément originale (13), a connu un processus général
d'extinction et s'est trouvée supplantée par une faune
quaternaire en très grande partie semblable (bien que notablement
appauvrie en ce qui concerne le nombre d'espèces) à
celle que nous connaissons aujourd'hui, c'est à dire très
proche de celle de l'Europe continentale voisine.
Deux points restent actuellement discutables : l'ancienneté
des peuplements de certaines espèces (en particulier le loir,
le hérisson et la belette) et l'importance de l'intervention
humaine dans l'établissement de l'ensemble de la faune Corse
actuelle (introductions volontaires ou accidentelles).
La thèse jusqu'ici accréditée privilégie
fortement l'hypothèse d'une faune mammalienne quasi- entièrement
introduite lors des premiers peuplements humains (actuellement datés
aux alentours de 7000 av. J.C.) et s'appuie essentiellement sur l'étude
des vestiges osseux disponibles (14).
Sans entrer dans la polémique portant sur la date effective
de l'arrivée des premiers hommes en Corse (15), quelques points
sont susceptibles de contredire en partie l'hypothèse présentée
ci-dessus :
- Du point de vue de l'importance des régressions marines successives
et de l'existence hypothétique de ponts continentaux, l'histoire
de la Méditerranée insulaire présente encore
des zones d'ombre.
- la rareté des vestiges osseux pléistocènes,
due en partie aux caractéristiques géologiques de la
Corse, rend difficile nombre de déterminations taxonomiques(1)
et ne permet pas d'affirmer que l'ensemble de la faune de cette époque
est définitivement connu.
1. On entend par taxonomie tout ce qui touche à l'identification et à la classification des organismes vivants.
- Il semble difficile d'imputer à l'homme l'introduction
d'espèces à priori peu commensales comme le loir, le renard
ou le chat sauvage, qui présentent, d'autre part, des caractéristiques
morphologiques marquées leur conférant, à tout
le moins, le statut de sous-espèce. Il faut signaler à
ce propos que la plupart des données permettant d'établir
des comparaisons entre peuplements insulaires et continentaux font actuellement
défaut et en particulier les caractérisations génétiques
(essentiellement basées sur les polymorphismes chromosomique
et protéique) qui sont le plus souvent utilisées pour
apprécier le degré de divergence entre populations ou
sous-espèces. Les rares travaux disponibles font état
d'une originalité incontestable (16) et confortent les observations
morphologiques qui ont souvent conduit à considérer nombre
de populations corses de Mammifères comme sensiblement différentes
de leurs homologues continentales.
- Le phénomène du marronnage (17) proposé pour
expliquer la présence du mouflon et du sanglier insulaires
est rendu problématique par les premières datations
des vestiges osseux de ces deux animaux en Corse et la date supposée
de leur domestication au Moyen-Orient (18). L'existence, pour le mouflon,
de populations comparables en Sardaigne (et d'une espèce fort
proche peuplant l'île de Chypre), n'est pas sans soulever un
certain nombre de questions : identité des peuplements, marronnages
systématiques, convergences phénotypiques… Relevons
également, en ce qui concerne le mouflon, que les arguments
génétiques avancés pour conforter la thèse
du marronnage (identité entre les variants de l'hémoglobine
du mouflon de Corse et du mouton domestique, coloration de type aberrant…)
sont extrêmement discutables, en particulier quant à
l'origine des animaux étudiés (19). Le cas du sanglier
est comparable à celui du mouflon et sa présence en
Corse sous une forme semi-domestique importée, aux dates indiquées
par les préhistoriens, paraît hautement improbable. Les
arguments cytogénétiques (un nombre de chromosomes,
2n = 38, identique à celui du porc domestique) ne semblent
pas en l'occurrence particulièrement déterminants pour
éclairer l'origine de cette espèce dans l'île
(20).
Ces remarques ne visent pas à substituer à une hypothèse
discutable une autre qui le serait tout autant, mais s'il semble évident
que pour bon nombre d'espèces l'intervention humaine est indéniable,
cela ne saurait constituer un argument suffisant pour rendre l'homme
responsable de la faune mammalienne corse actuelle.
L'hypothèse du "tout-introduit" peut même conduire
dans certains cas à des considérations erronées.
Ainsi la reconnaissance tardive de l'existence du chat sauvage de
Corse, bien qu'elle ait été depuis longtemps avancée
(21), est en grande partie due au scepticisme de la plupart des spécialistes
des peuplements insulaires quant à la possibilité d'existence
dans les îles méditerranéennes d'espèces
autochtones non-commensales. Le chat sauvage ne pouvait pas exister
en Corse, sauf sous forme de chat haret. En fait, au même titre
que le chat sauvage de Sardaigne, le ghjattu volpe existe bel et bien
(22). On ne peut que constater ici la pertinence de l'argument…
linguistique.
Signalons également, à propos des discussions que nourrit
tel ou tel problème biogéographique, le cas tout à
fait particulier de la martre (Martes martes), qui serait présente
dans l'île(2) mais dont l'existence est pour le moins hypothétique…(23).
(2) Cf. Noblet et al.(1987)
Nous ne reviendrons pas ici sur l'importance plus ou
moins grande de l'homme dans l'implantation des peuplements animaux
en Corse mais essayerons plutôt de dégager brièvement,
et dans un but prospectif, quelques considérations sur la faune
insulaire actuelle.
L'avenir de celle-ci apparaît évidemment liée
au sort des biotopes, eux-mêmes de plus en plus modifiés
ou menacés par les activités humaines. Une grande partie
du paysage corse a été, au cours des siècles,
anthropisé en profondeur par une activité agropastorale
omniprésente. Même si ces effets se sont atténués
dans la plupart des régions de l'île avec l'abandon des
territoires cultivés et leur retour à un stade végétal
de maquis plus ou moins haut (parfois de forêt), les incendies
pastoraux contribuent toujours fortement à la modification
du couvert végétal, surtout en plaine et en moyenne
montagne.
La Corse possède toutefois, si on la compare aux autres grandes
îles méditerranéennes, une couverture forestière
encore importante (24). Sans parler du problème toujours aigu
des incendies, l'avenir de la faune corse reste assujetti à
l'évolution de deux paramètres majeurs :
- la pression humaine, qui se fait essentiellement touristique, risque
assez rapidement de modifier les biotopes de manière d'autant
plus importante que ceux-ci sont plus fragiles (haute montagne, dunes
côtières, zones humides…) ou plus facilement "occupables"
(25).
- L'introduction d'animaux étrangers à l'île,
qu'il s'agisse d'espèces nouvelles ou de représentants
d'espèces autochtones (26), hypothèque le maintien de
la faune originale.
En ce qui concerne le deuxième point, la politique actuelle
de protection sanitaire est quasiment inexistante (27).
Mais, si l'aspect prophylactique est évidemment très
important, il faut également savoir que ces introductions menacent
l'équilibre génétique des populations locales
(28), particulièrement fragile en milieu insulaire. La condamnation
de telles pratiques est d'autant plus difficile que la spécificité
des populations locales est niée et/ou méconnue.
Le problème des introductions, surtout si elles intéressent
des espèces sauvages, semble relativement plus facile à
régler que celui de la dégradation de l'environnement,
mais encore faudra-t-il pour cela que la Corse se dote d'une législation
efficace dans ce domaine.
La gestion des biotopes corses (on parle aujourd'hui de gestion des
paysages…) soulève par contre un certain nombre d'interrogations
: comment résoudre au mieux l'équation mettant en relation
pêle-mêle le potentiel "sauvage" de l'île
(sous-entendu le potentiel touristique), la "désertification"
de l'intérieur (mais en fait de toutes les régions non-urbanisées),
la protection de zones encore préservées, le développement
touristique ?
Si la Corse ne se donne pas rapidement les moyens d'une politique
ambitieuse en matière d'environnement, elle risque de voir
se modifier de manière spectaculaire un paysage faunistique
déjà grandement dépendant des actions de protection
(29) et dont la gestion échappera, de plus en plus, à
la population insulaire elle-même.
La sauvegarde du patrimoine faunistique ne peut passer que par une
prise de conscience générale et, parallèlement
à cette problématique de la préservation et du
développement, ce sont les rapports même des Corses avec
la nature qui se doivent d'évoluer, en particulier pour tout
ce qui touche aux prélèvements halieutiques et cynégétiques
(30). Mais nous abordons là un domaine qui échappe au
cadre de la simple analyse biologiques…
(1) Cf. l'ancien mais remarquable travail de T. de Caraffa
(1929) sur le sujet.
(2) Même si c'est un facteur essentiel, la superficie d'une île
n'explique pas tout : il faut également considérer la
distance qui la sépare du continent le plus proche et la variété
de ses biotopes. Ainsi, la Corse, qui présente sensiblement la
même superficie que Chypre et la Crête, possède un
nombre d'espèces d'Oiseaux nettement supérieur grâce
à une diversité géographique plus grande. Cette
diversité la place presque au niveau de la Sardaigne pourtant
près de trois fois plus étendue mais nettement mois élevée
et dépourvue d'un certain nombre d'essences forestières
(sapin, hêtre, pin laricciu…) par rapport à elle.
(3) Ce sont la truite, l'anguille, la blennie fluviatile et l'épinoche.
L'origine de cette dernière, peut être introduite, est
discutée.
(4) C'est le cas des Fauvettes (Sylvia sp.), particulièrement
bien représentées dans le bassin méditerranéen
occidental.
(5) On entend par niche écologique tout ce qui est nécessaire
à la survie d'un organisme dans son biotope (ce qui sous-entend
aussi bien une notion d'ordre spatial qu'un système comportemental
bien précis).
(6) Le renard et la belette en sont des exemples flagrants. Leur éventail
de proies et de modes de prédation est d'autant plus large que
le nombre de carnivores de l'île est réduit.
(7) Dans les biotopes montagnards il atteint jusqu'à 60%, pour
certains taxons de la faune d'Invertébrés dulçaquicoles
(Guidicelli, 1968).
(8) Un certain nombre d'Oiseaux et de Mammifères présentent
toutefois des sous-espèces indiscutables, le plus souvent cyrnosardes.
Cf. pour détail Thibault (1983) et Noblet et al. (1987).
(9) Les caractéristiques génétiques des populations
de truites varient fortement d'une région de l'île à
l'autre. Elles reflètent sans doute aussi en partie la pollution
génétique due aux introductions à vocation piscicole
(Krieg et Guyomard, 1983).
(10) L'isolement géographique est à l'origine de la mise
en place des mécanismes d'isolement reproductif (Mayr, 1974 ;
Dobzhanski, 1980). Ceux-ci ont, en général, une efficacité
proportionnelle à la durée de cet isolement. Ils sont
dus en fait à la divergence génétique qui s'installe
progressivement entre deux populations isolées d'une même
espèce. En cas de remise en contact, les croisements entre individus
des deux populations seront impossibles (formation d'espèces
vraies, parfois qualifiées de "bonnes" espèces),
partiels (espèces "imparfaites" qu'on appelle espèces
vicariantes, vice species, semi species ou quasi species) ou bien normaux
(on parle alors simplement de populations ou de sous-espèces
si la divergence génétique est déjà sensible
et permet l'identification quasi-systématique des deux formes).
Il faut signaler la difficulté évidente d'établir
des définitions précises et fiables quant aux catégories
taxonomiques citées plus haut : en fait seule l'appréciation
du flux génique (c'est à dire concrètement du nombre
de croisements entre les individus de deux ensembles supposés
déjà plus ou moins isolés génétiquement)
constitue un argument biologique pertinent. Ce flux est supposé
décroître au fur et à mesure que l'identité
spécifique se précise. Les problèmes restent toutefois
posés en ce qui concerne la validité de la notion de sous-espèce
et des critères susceptibles d'être retenus pour sa définition:
morphologiques (y compris chromosomiques), physiologiques, biochimiques,
écologiques, éthologiques (comportementaux)… Les
recherches actuelles privilégient l'étude des polymorphismes
biochimiques à travers la mise en œuvre de différentes
techniques : les résultats sont ensuite traités mathématiquement
(fréquences géniques, taux d'hétérozygotes,
distances génétiques…) afin de préciser la
position d'une population ou d'une espèce par rapport à
d'autres. Signalons la part croissante que prend actuellement dans ce
type de travaux la génétique moléculaire basée,
elle, sur l'analyse "directe" des modifications du matériel
héréditaire premier, l'acide désoxyribonucléique
(ADN).
(11) Quatre espèces de perdrix sont réparties autour du
bassin méditerranéen : la rouge (Alectoris rufa) occupe
la péninsule ibérique, le sud de la France et l'Italie,
la bartavelle (Alectoris graeca) les Balkans, la chukar (Alectoris chukar)
le Moyen-Orient et la gambra (Alectoris barbara) l'Afrique du Nord.
On constate la présence d'hybrides dans les zones de contact
entre ces différentes espèces qui restent parapatriques
(cf. notes 10 et 12). Il est remarquable que les grandes îles
méditerranéennes possèdent une seule et unique
espèce de perdrix, différente pour chacune : présence
de la rouge en Corse (et également aux Baléares), de la
gambra en Sardaigne, de la bartavelle en Sicile et de la chukar à
Chypre. L'hypothèse de l'introduction humaine actuellement avancée
pourrait expliquer en partie cette répartition géographique.
En fait cette situation est surtout révélatrice de l'importance
des problèmes de compétition dans les biotopes insulaires
méditerranéens (cf. Blondel, 1986).
(12) En règle générale deux espèces ne peuvent
se partager une niche écologique identique. Il est possible que
dans certains cas la différenciation de la niche intervienne
plus ou moins tardivement par rapport aux mécanismes de spéciation.
Certaines espèces fort proches, occupant des aires géographiques
voisines (parapatriques), se côtoient sans se chevaucher : ce
sont des espèces dites vicariantes, en général
considérées comme des espèces récemment
formées et encore imparfaitement isolées reproductivement
(il existe de nombreux exemples pour la Corse et la Sardaigne). Cet
état de fait biologique explique l'inanité des efforts
d'introduction en Corse de différentes espèces de perdrix
allogènes (chukar et bartavelle en particulier).
(13) Au Pléistocène, la faune du massif cyrno-sarde est
fortement endémique et présente des espèces très
caractéristiques parmi lesquelles on peut citer le cerf de Caziot
(Megaceros cazioti), un carnivore très proche du chien (Cynotherium
sardous), une espèce de loutre marine (Nesolutra ichnusae), un
Primate du genre Macaca, une musaraigne géante (Episoriculus
corsicanus, un mulot et un campagnol géants ainsi qu'un éléphant
et un hippopotame nains. Toutes ces espèces vont disparaître
avant l'arrivée de l'homme néolithique sur l'île
à l'exception d'un étrange Lagomorphe, mi-lapin mi-rat,
le Prolagus sardus. Certaines populations de cette espèce encore
très abondante au Néolithique et à l'Age du Bronze
se sont maintenues jusqu'à l'époque historique, comme
celle de l'îlot sarde de Tavolara.
(14) Pour une vue exhaustive de ce sujet cf. l'importante thèse
de Vigne (1983) sur les origines du peuplement mammalien postglaciaire
de la Corse.
(15) La présence de l'homme en Sardaigne (formant à cette
époque avec la Corse le bloc cyrno-sarde) est attestée
au Paléolithique par les nombreux restes d'industrie lithique
de Laerru et Perfugas (Sassari). Leur classification chronologique précise
est encore difficile mais ils semblent bien devoir se rapprocher du
clactonien européen (450 000 à 150 000 ans av. J. C.).
Plus récents, les ossements de la grotte du Corbeddu, à
Oliena (Nuoro), mettent en évidence un peuplement humain remontant
au Paléolithique supérieur, soit 35 000 à 12 000
ans av.J.C. (Sondaar et al. 1984).
(16) C'est le cas pour le sanglier corse dont le caryotype est différent
de celui des populations d'Europe occidentale (2n = 38 chromosomes contre
2n = 36) et identique à celui du porc domestique (Franceschi,
1980 ; Popescu et al. 1980)
(17) C'est le retour à la vie sauvage d'animaux appartenant à
une espèce déjà domestiquée. Ce phénomène
touche aussi bien les porcins que les bovins, les caprins et les ovins.
Actuellement il concerne essentiellement, en Corse, l'espèce
caprine et quelques porcs, le plus souvent croisés, mais il ne
semble pas conduire à l'établissement de populations pérennes.
(18) Des restes d'Ovicaprinae ont été retrouvés
au niveau XVIII de l'abri d'Araguina Sennola, près de Bonifaziu
(Lanfranchi et Weiss, 1977). L'ancienneté de ces vestiges (environ
6 570 av. J.C.) en ferait la première trace connue de domestication
du mouton en Méditerranée occidentale. La taille de ces
animaux pose problème quant à leur origine : les premiers
moutons méditerranéens, tous plus récents, présentent
un format nettement inférieur à celui de la forme insulaire
(Mazzolini, 1988).
(19) Les arguments utilisés quant à l'origine par marronnage
du mouflon de Corse s'appuyant sur des études du polymorphisme
biochimiques et des mutations pigmentaires (Bunch et al. 1978 ; Denis
et al. 1979 ; Poplin, 1979) sont contestables : les mouflons étudiés
provenaient d'un parc zoologique où de nombreux croisements mouflon-mouton
domestique avaient eu lieu (cf. Battesti et al. 1992).
(20) De nombreuses sous-espèces de sanglier possèdent
un caryotype à 2n = 38 chromosomes (Afrique du Nord, Balkans,
Europe Centrale, Asie Centrale, Sibérie, Japon…). Alors
qu'on pensait le porc domestique (2n = 38) issu de sangliers dont le
caryotype à 2n =36 aurait subi un réarrangement chromosomique,
il semble de plus en plus certain que la domestication a en majorité
intéressé des populations d'animaux à 2n = 38.
(21) Lavauden a décrit comme une nouvelle espèce (Felis
reyi) un spécimen de chat sauvage Corse, capturé dans
les environs de Biguglia en 1929.
(22) On doit à Arrighi et Salotti (1988) d'avoir recueilli les
éléments qui ont permis la reconnaissance du chat sauvage
de Corse. Les caractéristiques de Félidé sont encore
à préciser, tant du point de vue biologique que taxonomique.
(23) Si elle peuple effectivement les grandes îles de la Méditerranée
occidentale (Baléares, Sardaigne, Sicile), la martre en Corse
n'est signalée que par quelques observateurs vraisemblablement
peu familiarisés avec les mammifères insulaires : la taille
très importante de certaines belettes corses et leur type de
coloration sont probablement à l'origine d'une confusion. On
peut toutefois envisager à partir de la Sardaigne (où
elle est cependant assez rare) et de l'île d'Elbe une hypothétique
colonisation de la Corse.
(24) La Corse possède près de 280 000 hectares de superficie
boisée dont, il est vrai, une part importante de maquis plus
ou moins haut.
(25) L'hémorragie démographique que subit l'intérieur
de l'île débouche sur des problèmes aigus de gestion
et de protection des paysages. L'abandon des activités agropastorales
traditionnelles laisse vacant un immense espace naturel qui a tendance,
du fait même de l'absence (ou du nombre trop réduit) de
ses utilisateurs et "régulateurs" légitimes,
à devenir au mieux un espace de totale liberté et au pire
une véritable zone de pillage : pillage touristique, certes,
mais également forte pression en mode épisodique de la
population insulaire citadine. Celle-ci, de plus en plus importante,
semble apparemment de moins en moins soucieuse ou tout simplement capable
de respecter un univers devenu fragile et dans lequel elle se reconnaît
d'autant plus difficilement qu'il apparaît comme le reflet d'une
société archaïque et dépassée, condamnée
à disparaître (1). L'ensemble de la faune (et de la flore)
souffre doublement de cet état de fait : aux prélèvements
immodérés des furesteri se greffe, de plus en plus souvent,
le braconnage intensif de paisani bien décidés à
profiter prioritairement de ressources dont ils s'estiment prioritairement
bénéficiaires (cf. également à ce sujet
la note 27).
1. Paradoxalement, la société Corse dite traditionnelle et le système de valeurs qu'elle véhicule, aujourd'hui proches de la disparition, servent toujours de point de référence et sont célébrés dans d'innombrables chansons et poésies.
(26) Cf. Battesti et al. (1992)
(27) Un rapport du CCECV (1985) met l'accent sur l'absence de contrôle
sérieux des vaccinations en ce qui concerne en particulier les
animaux de compagnie (chiens et chats) susceptibles d'introduire la
rage en Corse. Ceci reste valable pour les introductions d'animaux d'élevage
en provenance du continent qui continuent d'accroître le nombre
des maladies du bétail.
(28) A propos de l'adaptation génétique des populations
naturelles et du danger que représentent pour elles les introductions,
un des plus grands biologiste de notre époque est on ne peut
plus explicite :
"… le phénotype (1) de chaque population locale est
très précisément ajusté pour répondre
exactement aux exigences de l'environnement local. Cet ajustement est
le résultat d'une sélection de gènes produisant
le phénotype optimal. La découverte de cette adaptation
physiologique des populations locales est d'une importance pratique
considérable pour ce qui concerne par exemple le mode de vie
sauvage. Transportées dans différents autres environnements,
les populations qui sont bien adaptées à leur milieu natal
sont souvent très vulnérables. La littérature sur
le gibier rapporte nombre d'exemples où des animaux moururent
rapidement après avoir été transportés dans
une région différente. S'ils survivent suffisamment longtemps
pour s'accoupler, l'introduction de leurs gènes inférieurs
contribuera à la détérioration des populations
indigènes. C'est pour cette raison que certains pays interdisent
à présent l'importation de gibier avien et mammalien.
On aurait pu épargner des millions fournis par les contribuables
pour élever ou transplanter des souches de gibier mal adaptées
si les dirigeants avaient été au courant des différences
physiologiques des populations locales…" (Mayr, 1974).
1. Le phénotype est l'ensemble des caractéristiques apparentes d'un individu, c'est à dire le résultat de l'interaction entre l'ensemble de ses gènes (génotype) et l'environnement
(29) Un certain nombre d'espèces ont disparu au
cours des dernières décennies. Citons, parmi les plus
connues, le pygargue à queue blanche, l'érismature à
tête blanche, le cerf de Corse (réintroduit à partir
de la Sardaigne, en 1985, par le PNRC), le phoque moine. Il faut également
savoir que la survie des grands rapaces de l'île (aigle et surtout
gypaète) est de plus en plus subordonnée à l'action
humaine (en particulier nourrissage hivernale et surveillance des aires)
et que les espèces marines à effectifs réduits
(balbuzard et goéland d'Audouin principalement) sont menacées
par une pression touristique estivale croissante.
(30) Dalla Bernardina (1989), partant du mythe du paysan-écologiste,
analyse la perception apocalyptique de la disparition du gibier qu'ont
les Corses et qu'ils s'interprètent comme le résultat
du progrès et d'une intervention étrangère à
leur contexte culturel.
ARRIGHI j., SALOTTI M., 1988 : Le chat sauvage (Felis sylvestris schreber, 1777) en Corse. Confirmation de sa présence et approche taxonomique. Mammalia, 52, 123-135.
BATTESTI P., FRANCESCHI P., SALOTTI M., 1992 : les introductions de Mammifères sauvages en Corse. Introductions et réintroductions de Mammifères sauvages. XIVème colloque francophones de Mammalogie de la SFEPM, Orléans, Octobre, 1990. Nature centre. Annales biologiques du centre, 231-245.
BLONDEL J., 1982 : Caractérisation et mise en place des avifaunes dans le bassin Méditerranéen, Ecologia méditerranea, 8,253-272.
BLONDEL J., 1986 : biogéographie évolutive. Masson ed. , Paris.
BUNCH T.D., N'GUYEN T.C., LAUVERGNE J.J., 1978 : Hemoglobins of the Corsico-Sardinian Mouflon (ovis musimon) and their implications for the origin of HbA in the domestic sheep (Ovis aries) Ann. Génét. Sél. Anim. , 10, 503-506.
CARAFFA (de) T., 1929 : Les poissons de mer et la pêche sur les côtes de la Corse. Laffitte Reprints Ed., Marseille.
Conseil de la Culture de l'Education et du Cadre de Vie (1985) : la rage en Corse. Rapport dactylographié, Assemblée de Corse, Ajaccio, 6 p.
DALLA BERNARDINA S., 1989 : La disparition du gibier comme métaphore de l'apocalypse : un exemple Corse. L'île-miroir. Centre d'Etudes Corses. Acta Colloque Aix-en-Provence, novembre 1987. La Marge Ed., Ajaccio. 59-72.
DELAUGERRE M., CHEYLAN M., 1992 : Atlas de répartition des Batraciens et Reptiles de Corse. PNRC et Ecole Pratique des Hautes Etudes Ed., Ajaccio.
DENIS B., LAUVERGNE J.J., THERET M., 1978 : Un variant clair du Mouflon Corsico-Sarde (Ovis musimon) dû à un allèle au locus B (Brun). Ann. Génét. Sél. anim. , 10, 507-516.
FRANCESCHI P., 1980 : Essai de caractérisation génétique du porc corse. Aspects cytogénétiques et polymorphisme biochimique. Thèse Paris VI.
GUIDICELLI J., 1968 : Recherches sur le peuplement, l'écologie et la biogéographie d'un réseau de la Corse centrale. Thèse Marseille.
KRIEG F., GUYOMARD R., 1983 : Mise en évidence électrophorétique d'une forte différence génétique entre populations de Truite Fario de Corse. C.R. Acad. Sc. Paris, 296, 1089-1094.
LANFRANCHI (de) F., WEISS M.C., 1977 : Araguina-Sennola, dix années de fouilles préhistoriques à Bonifacio. Archeologia Corsa, 2, 1-167.
LAVAUDEN L., 1929 : Sur le chat sauvage de la Corse. C.R.Acad. Sc. Paris, 189,1023-1024.
MAYR E., 1974 : Populations, espèces et évolution. Herman Ed., Paris.
MUZZOLINI A., 1988 : Une ébauche de scénario pour le peuplement ovin ancien dans le Bassin méditerranéen. Populations traditionnelles et premières races standardisées d'Ovicaprinae. Gontard, Juillet 1987. Les colloques de l'INRA, 47, 289-298.
NOBLET J.F., CHEYLAN G., DEMEAUTIS G., DUBRAY D., FONS R., FRANCESCHI ., VIALE D., GAUTHIER A., ORSINI P., SALOTTI M., VIGNE J.D., 1987 : Les Mammifères en Corse. Espèces éteintes et actuelles. PNRC, Ajaccio.
POPESCU C.P., QUERE J.P., FRANCESCHI P.F., 1980/ Observations chromosomiques chez le sanglier français. Ann. Génét. Sél. Anim. , 12, 395-400.
POPLIN F., 1979 : Origine du Mouflon de Corse dans une
nouvelle perspective : par marronnage. Ann. Génét. Sél.
Anim. , 11, 133-143.
SONDAAR P.Y., BOER P.L., SANGES M., KOTSAKIS T., ESU D., 1984 : First
report on a Paleolithic culture in Sardinia. BAR, Int. Séries,
229, 29-47.
THIBAULT J. C., 1983 : Les Oiseaux de la Corse Histoire et répartition aux XIXe et Xxe siècles. PNRC, Ajaccio.
VIGNE J.D., 1983 : Les Mammifères non-volants du Post-Glaciaire de Corse et leurs rapports avec l'Homme : étude paléo-ethno-zoologique fondée sur les ossements. Thèse Paris.